BET PROTAC
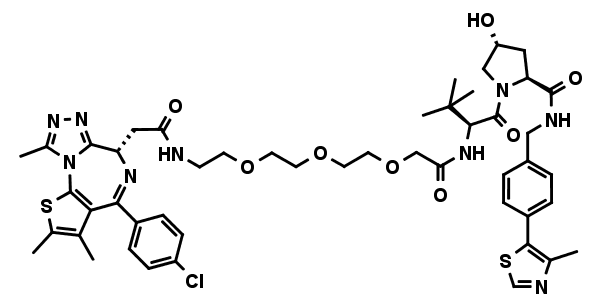
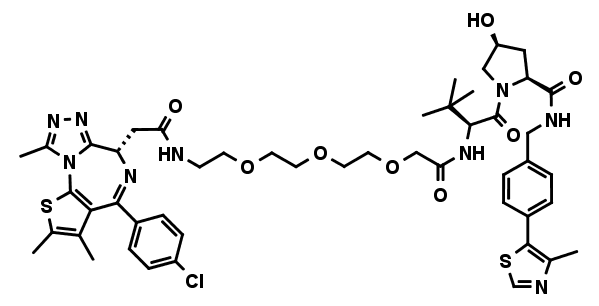
MZ1
MZ1 is a first in class PROTAC (proteolysis-targeting chimeras) that tethers JQ1 to a VHL E3 ubiquitin ligase ligand, aimed at triggering the intracellular destruction of BET proteins. It induces reversible, long-lasting, and preferential removal of BRD4 over BRD2 and BRD3 in cells. MZ1 and the negative control cis MZ1 have been discovered by Michael Zengerle, Kwok-Ho Chan, and Alessio Ciulli1 at the Ciulli Lab at the University of Dundee, School of Life Sciences. We are grateful to Alessio Ciulli for making this molecule available for scientific research via opnMe.



More information
Acetylation of lysine residues is a post-translational modification with broad relevance to cellular signalling and disease biology. Inhibitors that modulate the ‘reading process’ mediated by acetyl lysines are an area of extensive research. The principal readers of ɛ-N-acetyl lysine (Kac) marks are bromodomains (BRDs), which are a diverse family of evolutionary conserved protein-interaction modules. The conserved BRD fold contains a deep, largely hydrophobic acetyl lysine binding site, which represents an attractive pocket for the development of small, pharmaceutically active molecules. Proteins that contain BRDs have been implicated in the development of a large variety of diseases.
Traditionally small molecules have been used to inhibit the action of a target protein by occupying and blocking a functional region of the protein. An alternative innovative approach is the development of proteolysis targeting chimeras (PROTACs), i.e. heterobifunctional compounds consisting of one moiety that binds a Cullin RING E3 ubiquitin ligase linked to another that binds a desired protein of interest (POI), bringing the ligase and the POI into close spatial proximity. This hijacks the intrinsic catalytic activity of the E3 ligase from the natural and directs it toward the POI as a neo-substrate, triggering its poly-ubiquitination and subsequent proteasome-dependent degradation. As a result, a PROTAC acts as a degrader of the target as opposed to just an inhibitor, enabling the effective post-translational elimination of a target gene product in living organisms. This approach presents many advantages compared to conventional target inhibition. One of the most attractive features of the approach is that a PROTAC molecule acts sub-stoichiometrically, i.e. it only needs to bind a molecule of target once to induce its degradation, and then is released and set free to bind another molecule of target and carry on, as in a catalytic cycle. For this reason, the concentrations required for PROTACs to be active in cells tend to be much lower compared to those needed to be reached and maintained with inhibitors, which can lead to fewer off-target effects and a more selective chemical intervention on the desired target.
Structural elucidation of the key ternary ligase-PROTAC-target species and its impact on target degradation selectivity remain elusive. The crystal structure of BRD4 degrader MZ1 in complex with human VHL and the BRD4 bromodomain (BRD4BD2) offers unique insights into the first ternary complex structure of a low molecular weight degrader. The ligand folds into itself to allow formation of specific intermolecular interactions in the ternary complex. Isothermal titration calorimetry studies, supported by surface mutagenesis and proximity assays, are consistent with pronounced cooperative formation of ternary complexes with BRD4BD2. Our results elucidate how PROTAC-induced de novo contacts dictate preferential recruitment of a target protein into a stable and cooperative complex with an E3 ligase for selective degradation1,2.
Researchers can request up to two batches of 5 mg of MZ1 and one batch of cis MZ1 for free. Larger quantities of the compounds are available from Tocris.
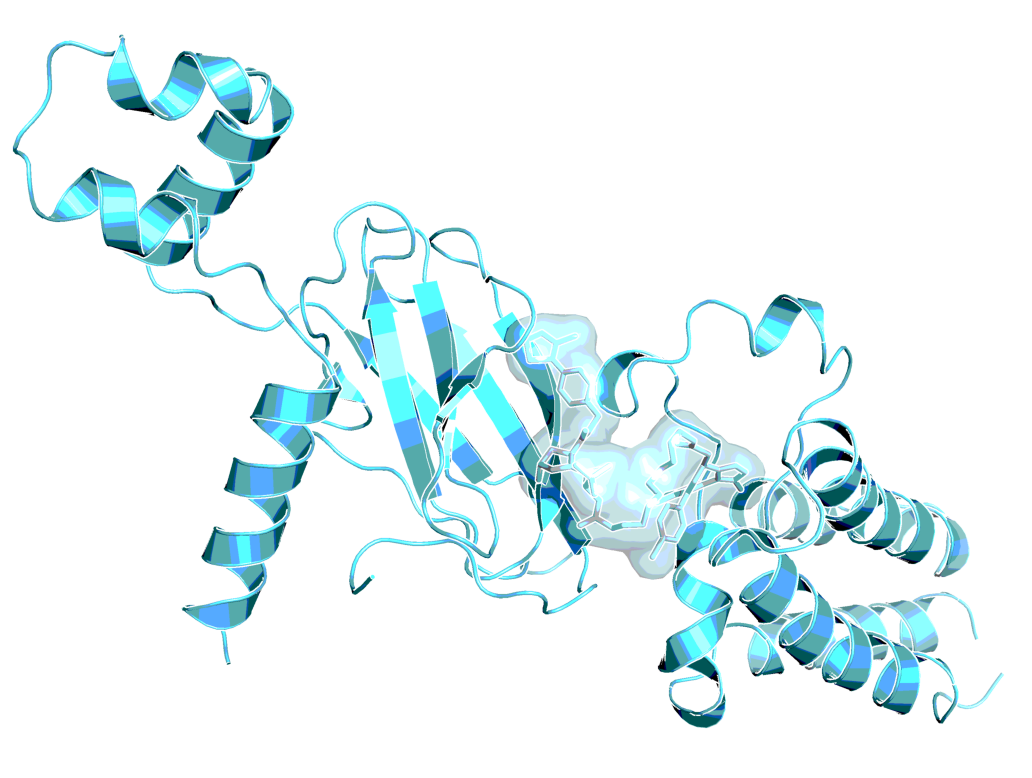
MZ1, 3D conformation, as observed in complex with BRD4 bromodomain and VHL (PDB 5T35)
The binary affinity for MZ1 binding to BRD4BD2 and to VCB (VHL-ElonginC-ElonginB) is 15 and 66 nM respectively (ITC). The affinity of the negative control “cis MZ1” to VCB is > 15 µM (ITC). In presence of excess of BRD4BD2 MZ1 the ternary KD for the BRD4BD2::MZ1::VCB is 3.7 nM. MZ1 degrades the BRD4 protein with a DC50 of 2-20 nM depending on the cell line, and with selectivity on DC50 of ~10-fold relative to BRD2 and BRD3.
Table: Binding affinities for binary and ternary complexes between MZ1, VCB and BET bromodomains as measured by isothermal titration calorimetry (for assay conditions/details see reference 2).
| PROBE NAME | MZ1 | ||
| Protein in syringe | Species in cell | Kd (nM) | α | Δp<Kd |
| BRD2BD1 | MZ1 | 62 ± 6 | - | - |
| BRD2BD2 | MZ1 | 60 ± 3 | - | - |
| BRD3BD1 | MZ1 | 21 ± 5 | - | - |
| BRD3BD2 | MZ1 | 13 ± 3 | - | - |
| BRD4BD1 | MZ1 | 39 ± 9 | - | - |
| BRD4BD2 | MZ1 | 15 ± 1 | - | - |
| BRD2BD1 KEA | MZ1 | 69 ± 9 | - | - |
| BRD4BD1 QVK | MZ1 | 22 ± 8 | - | - |
| VCBa | MZ1a | 66 ± 6 | - | - |
| VCB | MZ1:BRD2BD1 | 24 ± 8 | 2.9 | 0.4 ± 0.2 |
| VCB | MZ1:BRD2BD2 | 28 ± 3 | 2.3 | 0.36 ± 0.06 |
| VCB | MZ1:BRD3BD1 | 19 ± 4 | 3.5 | 0.5 ± 0.1 |
| VCB | MZ1:BRD3BD2 | 7 ± 2 | 10.7 | 1.0 ± 0.2 |
| VCB | MZ1:BRD4BD1 | 28 ± 6 | 2.3 | 0.4 ± 0.1 |
| VCB | MZ1:BRD4BD2 | 3.7 ± 0.7 | 17.6 | 1.24 ± 0.09 |
| VCB | MZ1:BRD2BD1 KEA | 12 ± 7 | 7.9 | 0.8 ± 0.3 |
| VCB | MZ1:BRD4BD2 QVK | 14.9 ± 0.1 | 4.2 | 0.62 ± 0.04 |
All ITC titrations were performed at 25 °C. Values reported are the mean ± S.E.M. from two independent measurements, except for VCB titration into MZ1 (line a) for which values reported are the mean ± S.E.M. from eight independent measurements.
The binding thermodynamics and kinetics of ternary complex formation and dissociation of MZ1 with VHL and the various BET bromodomains was also later investigated by the Ciulli Lab, in collaboration with Boehringer Ingelheim, using surface plasmon resonance (SPR). The data confirmed MZ1 forms the most stable and cooperative complexes between VHL and the second bromodomains (BD2s) of BET proteins. Specifically, the most stable, high-affinity and long-lived complex was formed with BRD4BD2, with a single residue (Gly386) contributing to this “glueing” in an isoform-specific manner, as demonstrated by data obtained with reciprocal swap Gly-Glu mutants in BRD3BD2 and BRD4BD2 4.
Table: Binding of MZ1 (binary complex formation) or MZ1:BET bromodomains (ternary complex formation) to surface-immobilized VCB as measured by surface plasmon resonance (for assay conditions/details see reference 4).
| SPR BINDING TO VHL | ||||
| PROTAC | +bromodomain target | Kd (nM) | α | t1/2 (s) |
| MZ1 | - (binary) | 29 | - | - |
| MZ1 | BRD2BD1 (ternary) | 23 | 1.3 | <0.7 |
| MZ1 | BRD2BD2 (ternary) | 0.9 | 32 | 67.4 |
| MZ1 | BRD3BD1 (ternary) | 12 | 2.4 | <0.7 |
| MZ1 | BRD3BD2 (ternary) | 8 | 3.6 | 6 |
| MZ1 | BRD3BD2 E344G (ternary) | 2 | 14 | 58.7 |
| MZ1 | BRD4BD1 (ternary) | 30 | 0.9 | <0.7 |
| MZ1 | BRD4BD2 (ternary) | 1 | 22 | 130 |
| MZ1 | BRD4BD2 G386E (ternary) | 8 | 3.5 | 4 |
MZ1 is soluble at physiological pH, metabolic stable (hepatocytes) and a PGP substrate (high Caco2 efflux ratio. Therefore, MZ1 is not suitable for oral administration but suitable for parenteral administration (i.v., i.p. or s.c.). The compound shows moderate inhibition of CYP3A4 and species differences in plasma protein binding.
| PROBE NAME / NEGATIVE CONTROL | MZ1 | CIS MZ1 |
| MW [Da, free base]a | 1,002.6 | 1,002.6 |
| logD @ pH 11 | 3.6 | 3.7 |
| Solubility @ pH 6.8 [µg/mL] | 19 | 12 |
| Caco-2 permeability AB @ pH 7.4 [*10-6 cm/s] | <0.34 | 0.3 |
| Caco-2 efflux ratio | 481 | 127 |
| Microsomal stability (human/mouse/rat) [% QH] | 88 / 79 / 73 | >95 / >95 / 71 |
| Hepatocyte stability 50% Serum (human/mouse/rat) [% QH] | 12 / 19 / 58 | n.d. |
| Plasma Protein Binding (human/mouse/rat/10%FCS) [%fu] | 2.5 / 0.42 / 2.5 / 41 | n.d. |
| CYP 3A4 (IC50)[µM] | 5.5 | n.d. |
| CYP 2C8 (IC50)[µM] | 24 | n.d. |
| CYP 2C9 (IC50)[µM] | >50 | n.d. |
| CYP 2C19 (IC50)[µM] | >50 | n.d. |
| CYP 2D6 (IC50)[µM] | 37 | n.d. |
a For the salt form you will get, please refer to the label on the vial and for the molecular weight of the salt, please refer to the FAQs
MZ1 is suitable for a parenteral administration (i.v., i.p. or s.c.). The compound shows high clearance in rats and low clearance in mice. High AUC levels can be obtained, when the compound is administered subcutaneously using a 25% HP-β-CD formulation. Because of the high Caco-2 efflux ratio, the oral exposure is very low.
| MZ1 | RATA | MOUSEB |
| Clearance [% QH] | 156 | 20.7 |
| Mean residence time after i.v. dose [h] | 0.61 | 0.34 |
| t1/2,term [h] | 1.52 | 1.04 |
| Vss [L/kg] | 4.05 | 0.38 |
| AUC0-inf [nM.h] | 1,300 | 3,760 |
| cmax [nM] | 163 | 2,070 |
| tmax [h] | 2 | 0.5 |
| t1/2,term [h] | 5.13 | 2.95 |
| cmax [nM] | - | <2 |
| F [%] | n.a. | 83 |
a rat i.v. dose: 5 mg/kg; rat s.c. dose: 5 mg/kg
b mouse i.v. dose: 5 mg/kg, mouse s.c. dose: 5 mg/kg
MZ1 shows antiproliferative and Myc-suppression activity in AML MV4;11 and HL60 cells. BET Reduction by MZ1 in HeLa Cells, Antiproliferative Activity, and BRD4/cMyc Reduction in AML Cellsa3.
MZ1 | |||||||||||
pDC50/Dmax (%) in HeLa cells | pIC50 | Emax (%) | BRD4/cMyc depletion (%) | (α) | |||||||
BRD4 | BRD4 | BRD3 | BRD2 | MV4;11 | HL60 | MV4;11 | HL60 | MV4;11 | HL60 |
| |
| MZ1 | 8.1 / 98 | 8.6 / 100 | 7.0 / 100 | 7.4 / 98 | 7.57 ± 0.03 | 6.66 ± 0.05 | 96.1 ± 0.3 | 92.1 ± 0.4 | 96 / 84 | 82 / 68 | 7.4 |
aDC50: concentration in molar causing 50% reduction of protein level relative to vehicle control treatment in 24h. Dmax: maximum reduction of protein level relative to vehicle control treatment. pIC50 was measured after 48h treatment. Errors on pIC50 values reflect the quality of the curve fitting. Protein depletion % are for 50 nM treatments (4h) in MV4;11/HL60. For more details please refer to reference 3.
“Cis MZ1” is the (S) hydroxy diastereoisomer of MZ1. While exhibiting MZ1 comparable Bromodomain binding affinity it no longer is able to bind and recruit VCB and therefore is not degrading BET proteins in cells. The affinity of the “cis MZ1” to VCB is > 15 µM (ITC).
.png)
“cis MZ1” which serves as a negative control
| SELECTIVITY DATA AVILABLE | MZ1 | CIS MZ1 |
| SafetyScreen44™ with kind support of | Yes | Yes |
| Invitrogen® | No | No |
| DiscoverX® | No | No |
| Dundee | No | No |
To assess the cellular selectivity of MZ1 for BRD4/3/2 degradation and identify potential degradation off-targets, multiplexed isobaric tagging mass spectrometry proteomic experiments were performed by the Ciulli Lab to monitor protein levels in a quantitative and unbiased manner. HeLa cells were treated in duplicate with DMSO, 1 mM MZ1, or 1 mM cis MZ1 for 24 h. Among the 5,674 proteins quantified in this analysis, of those that met the criteria for a statistically significant change in abundance, markedly selective degradation of BET proteins was observed, and in particular preferential degradation of BRD4 over BRD3 and BRD2. As expected, BRD4 was not depleted by treatment with negative control cis MZ1. Protein levels of other bromodomain-containing proteins remained unaffected2.
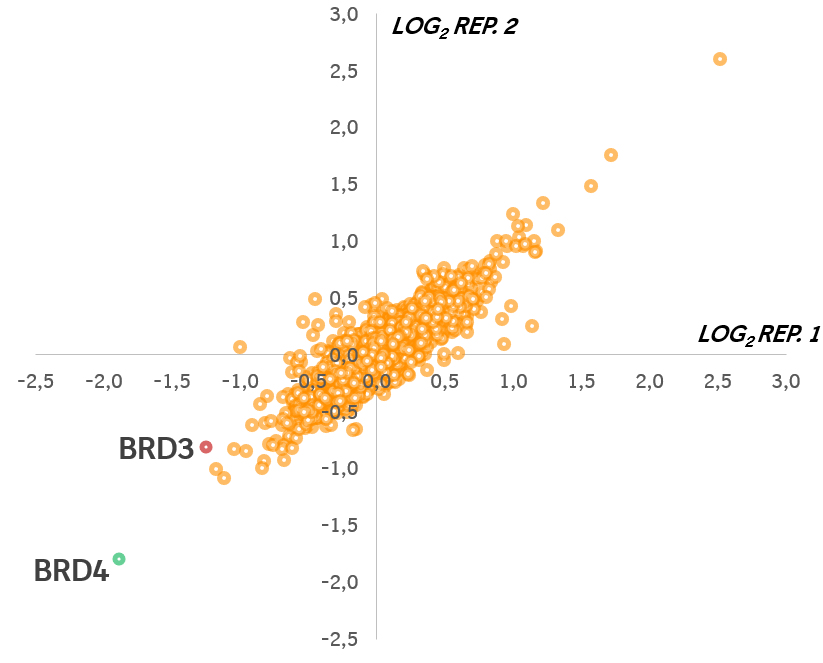
Impact of MZ1 (1 μM, 24 h) on the cellular proteome using global protein quantification by TMT-labeling mass spectrometry. Data plotted as fold change (log2) of replicate 1 versus replicate 2, for a total of 5,674 proteins quantified (for assay conditions/details see reference 2).
The PROTAC MZ1 was also tested by the Eric Fischer Laboratory - Dana-Farber Cancer Institute as part of their Degradation Proteomics Initiative5,6. It induces selective degradation of BRD2, BRD3 and BRD4 after 5 h of treatment at 1 µM in the neuroblastoma cell line Kelly Cells.
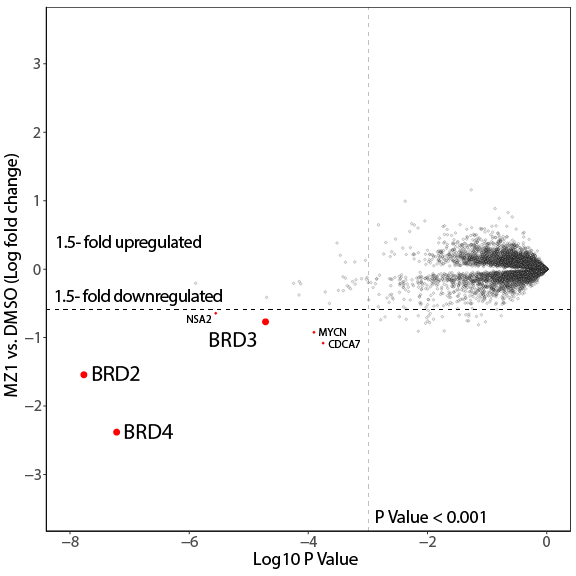
Global protein quantification was used to explore the unbiased proteome-wide selectivity of MZ1 induced degradation. Whole cell protein quantification was performed using label free quantification with the Fischer lab’s diaPASEF workflow.
Of the 7,084 proteins quantified in this experiment after in-house filtering for data quality, Donovan et al. found a high degree proteome-wide selectivity for BRD2, BRD3 and BRD4, confirming preferential degradation of BRD4, over BRD2 and BRD3. Statistical analysis was performed comparing degrader treatment to DMSO vehicle control using a moderated t-test in Bioconductor’s limma package to generate hit lists containing log2 Fold Change and P-values for each protein.
Download selectivity data:
MZ_1_selectivityData_0.xlsx
cis_MZ_1_selectivityData_0.xlsx
The X-ray of the PROTAC MZ1 in complex with the second bromodomain of BRD4 and pVHL:ElonginC:ElonginB is available in the PDB: 5T352.
dBET (CRBN based PROTAC) is a BET bromodomain degrader which is less specific than MZ17,8.
MZ1 is a BET protein family-specific PROTAC that originated from Alessio Ciulli’s lab at the University of Dundee, School of Life Sciences1.
{kind=link}
{kind=link}
Impact of Target Warhead and Linkage Vector on Inducing Protein Degradation: Comparison of Bromodomain and Extra-Terminal (BET) Degraders Derived from Triazolodiazepine (JQ1) and Tetrahydroquinoline (I-BET726) BET Inhibitor Scaffolds
Chan K.-H., Zengerle M., Testa A., Ciulli A.
J. Med. Chem. 2018, 61, 504-513.
The data can be accessed via the following link:
Mapping the Degradable Kinome Provides a Resource for Expedited Degrader Development
Donovan K. A., Ferguson F. M., Bushman J. W., Eleuteri N. A., Bhunia D., Ryu S., Tan L., Shi K., Yue H., Liu X., Dobrovolsky D., Jiang B., Wang J., Hao M., You I., Teng M., Liang Y., Hatcher J., Li Z., Manz T. D., Groendyke B., Hu W., Nam Y., Sengupta S., Cho H., Shin I., Agius M. P., Ghobrial I. M., Ma M. W., Che J., Buhrlage S. J., Sim T., Gray N. S., Fischer E. S.
Cell. 2020, 183(6):1714-1731.
When you plan a publication, please use the following acknowledgement:
MZ1 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.