BRD7/BRD9 PROTAC
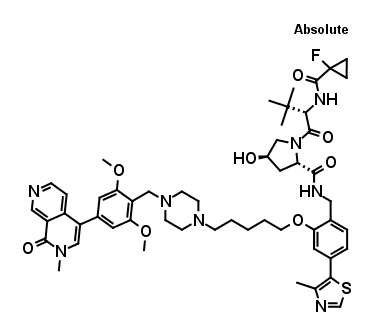
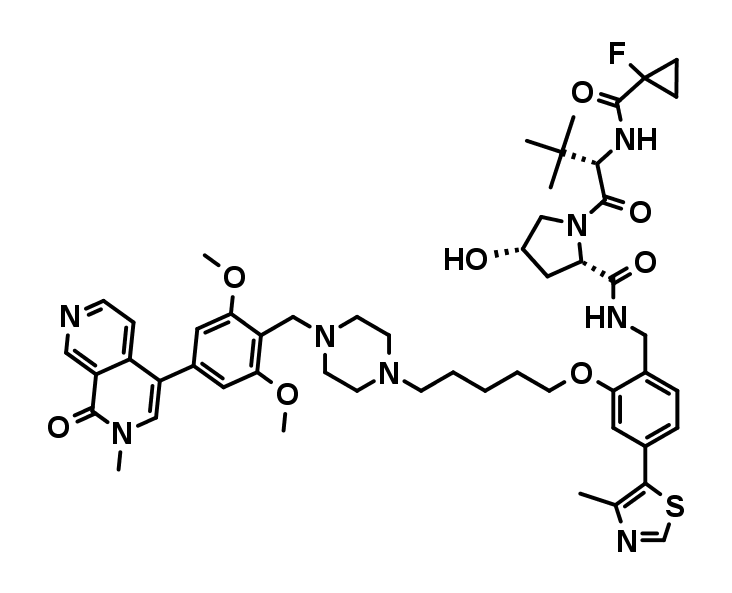
VZ185
VZ185 is a potent, fast, and selective degrader of BRD9 and its close homolog BRD7. It has shown a good in vivo PK profile, with high stability in plasma and microsomes as well as high aqueous kinetic solubility (up to ~100 μM). This compound is suitable for both in vitro and in vivo studies. The molecule and its negative control cis VZ185 have been discovered by Vittoria Zoppi, Scott Hughes, Chiara Maniaci, Andrea Testa, and Alessio Ciulli at the University of Dundee, School of Life Sciences, in collaboration with researchers from Boehringer Ingelheim and Promega.




More information
Developing PROTACs (proteolysis-targeting chimera) to redirect the ubiquitination activity of E3 ligases and potently degrade a target protein within cells can be a lengthy and unpredictable process, and it remains unclear whether any combination of E3 and target might be productive for degradation. We describe a probe-quality degrader for a ligase−target pair deemed unsuitable: the von Hippel−Lindau (VHL) and BRD9, a bromodomain-containing subunit of the SWI/SNF chromatin remodeling complex BAF. VHL-based degraders could be optimized from suboptimal compounds in two rounds by systematically varying conjugation patterns and linkers and monitoring cellular degradation activities, kinetic profiles, and ubiquitination, as well as ternary complex formation thermodynamics. The emerged structure−activity relationships guided the discovery of VZ185, a potent, fast, and selective degrader of BRD9 and of its close homolog BRD7. Our findings qualify a new chemical tool for BRD7/9 knockdown and provide a roadmap for PROTAC development against seemingly incompatible target−ligase combinations1.
BRD9 and its close homolog BRD7 (85% sequence identity2) are bromodomain-containing subunits of the BAF (BRG-/BRM-associated factor) and PBAF (Polybromo-associated BAF) complexes, respectively3,4. BAF and PBAF represent two variants of the SWI/SNF complex, one of the four mammalian ATP-dependent chromatin remodeling complexes. The SWI/SNF complexes control gene expression, DNA replication and DNA repair by modulating access to promoters and coding regions of DNA through modification of the degree of compactness of chromatin5-7. Mounting evidence from genetics and sequencing of cancer-associated mutations have spurred efforts to unravel yet largely elusive physiological roles of BAF/PBAF subunits and to develop targeted therapeutics in cancer and other human diseases3. In particular, BRD9 is overexpressed in several malignancies, such as cervical cancer and in non-small cell lung cancer (NSCLC)8,9. In contrast, BRD7 gene has been proposed as candidate tumor suppressor gene,10–13 as it regulates breast cancer cell metabolism14, and acts as negative regulator of aerobic glycolysis essential for tumor progression15. BRD7 also promotes X-box binding protein 1 (XBP1) nuclear translocation, which prevents the development of insulin-resistance disorders16. In contrast to these roles, it has been recently shown that inactivation of the BRD7 gene sensitizes tumor cells to T cell-mediated killing, suggesting that knockdown of BRD7 could be an attractive target for cancer immunotherapy17. Potent and selective inhibitors that bind to the BRD7/9 bromodomains have recently emerged from structure-guided medicinal chemistry campaigns, including compounds I-BRD9,2 LP99,18 ketone “compound 28”19, BI-7273 and BI-9564,1,20 and GNE-37521. These BRD7/9 inhibitors have been used in cells to help clarify the roles of the BRD7/9 bromodomain in oncogenesis and other disease states. For example, pharmacological studies of inhibitors BI-7273 and BI-9564 in combination with domain-swap protein engineering revealed that an active bromodomain of BRD9 is required to sustain MYC transcription and proliferation of leukemic cells20,22. These findings and availability of bromodomain ligands prompted us to initiate a PROTAC medicinal chemistry campaign to target BRD7 and BRD9 proteins for degradation1. Larger quantities of the compounds are available from Tocris.

Cocrystal structure of BRD9-BD and compound 5 as shown in reference 1 (PDB code 5EUI)
VZ185 displays binary and ternary KD = 30 nM to VHL in both ITC and FP binding assays and degrades BRD9 and BRD7 proteins with DC50 values of 1.8 nM and 4.5 nM and Dmax of 95% (in RI-1 cell lines)1. The total stability ΔG of the ternary complex VHL:VZ185:BRD9-BD is –21.7 kcal/mol1.
| PROBE NAME / NEGATIVE CONTROL | VZ185 | cis-VZ185 |
| MW [Da, free base]a | 995.2 | 995.2 |
| ITC (VHL binary KD) [nM]b | 26 ± 9 | n.d. |
| ITC (VHL ternary KD i.e. in the presence of BRD9-BD) [nM]b | 27 ± 3 | n.d. |
| ITC (BRD9-BD binary KD) [nM]b | 5.1 ± 0.6 | n.d. |
| ITC total ΔG (kcal mol-1)b | –21.7 | n.d. |
| FP (VHL binary KD) [nM]b | 35 ± 5 | n.d. |
| FP (VHL ternary KD i.e. in the presence of BRD9-BD) [nM]b | 35 ± 6 | n.d. |
| ITC/FP (Cooperativity, α)b | 1.0 | n.d. |
| Western Blot degradation assay (DC50, 8 h in RI-1 cells, BRD9 / BRD7) [nM]b | 1.8 / 4.5 | n.d. |
| Live-cell degradation (DC50, in HEK293 cells, HiBiT-Brd9 / HiBiT-Brd7) [nM]b | 4.0 / 34.5 | n.d. |
| WES degradation assay (DC50, 18 h BRD9, EOL-1 / A204 cells) [nM]b | 2.3 / 8.3 | n.d. |
| CellTiterGlo (Cell viability EC50, EOL-1 / A204 cells) [nM]b | 3.4 / 39.8 | n.d. |
a For the salt form you will get, please refer to the label on the vial and for the molecular weight of the salt, please refer to the FAQs
b Assay conditions available in reference 1
In vitro PK data further showed high stabilities of VZ185 in both plasma and microsomes from both human and mouse species, as well as high aqueous kinetic solubility (up to ∼100 μM). Together, the data qualify VZ185 as a novel high-quality degrader probe for cellular and potentially in vivo investigations.
| PROBE NAME / NEGATIVE CONTROL | VZ185 | CIS-VZ185 |
| logD @ pH 11 | 4.1 | 4.2 |
| CHI LogD @ pH 7.4a | 2.3 | 2.4 |
| Solubility (nephelometry) [µM]a | 85 | 79 |
| PAMPA permeability @ pH 7.4 Pe [nm/s]a | 0.01 | 0.36 |
| PAMPA permeability @ pH 7.4 recovery [%]a | 70 | 87 |
| Microsomal stability (human / mouse) [mL/min/g liver]a | 3.8 / 1.2 | 8.1 / 2.4 |
| Plasma stability T 1/2(human / mouse) [min]a | <180 / <180 | <180 / <180 |
a Assay conditions available in reference 1
| VZ185 | MOUSE | RAT |
| Clearance [% QH]a | 12 | 120 |
| Mean residence time after i.v. dose [h]a | 2.1 | 2.1 |
| tmax [h]b | 0.5 | 1.7 |
| F [%]b | Quantitatively bioavailable | Quantitatively bioavailable |
| Vss [L/kg] | 1.3 | 10 |
| AUCinfa [h*nmol/L] | 7,800 | 1,000 |
| AUCinfb [h*nmol/L] | 7,400 | 1,600 |
| AUDa [h*nmol/L] | 7,800 | 1,000 |
| AUDa [h*nmol/L] | 6,700 | 1,600 |
a i.v. dose: 5 mg/kg
b s.c. dose: 5 mg/kg
cis-VZ185 is the (S) hydroxy diastereoisomer of VZ185. While exhibiting comparable bromodomain binding affinity it no longer is able to bind and recruit the E3 ligase VHL and therefore does not induce the degradation of BRD7 and BRD9 proteins in cells.
cis VZ185 which serves as a negative control
| SELECTIVITY DATA AVAILABLE | VZ185 | cis-VZ185 |
| SafetyScreen44™ with kind support of | Yes | Yes |
| Invitrogen® | No | No |
| DiscoverX® | No | No |
| Dundee | No | No |
To assess the cellular selectivity of VZ185 for BRD7/9 degradation and identify potential degradation off-targets, multiplexed isobaric tagging mass spectrometry proteomic experiments were performed to monitor protein levels in a quantitative and unbiased manner.
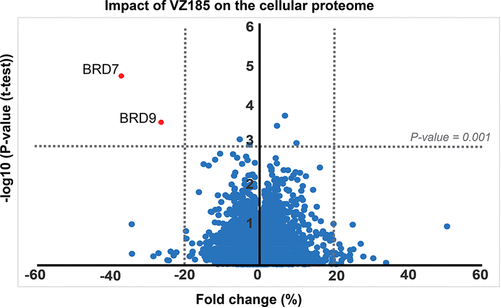
RI-1 cells were treated in triplicate with DMSO, 100 nM VZ185, or 100 nM cis VZ185 for 4 h. Among the 6,273 proteins quantified in this analysis, of those that met the criteria for a statistically significant change in abundance, markedly selective degradation of BRD7 and BRD9 was observed.
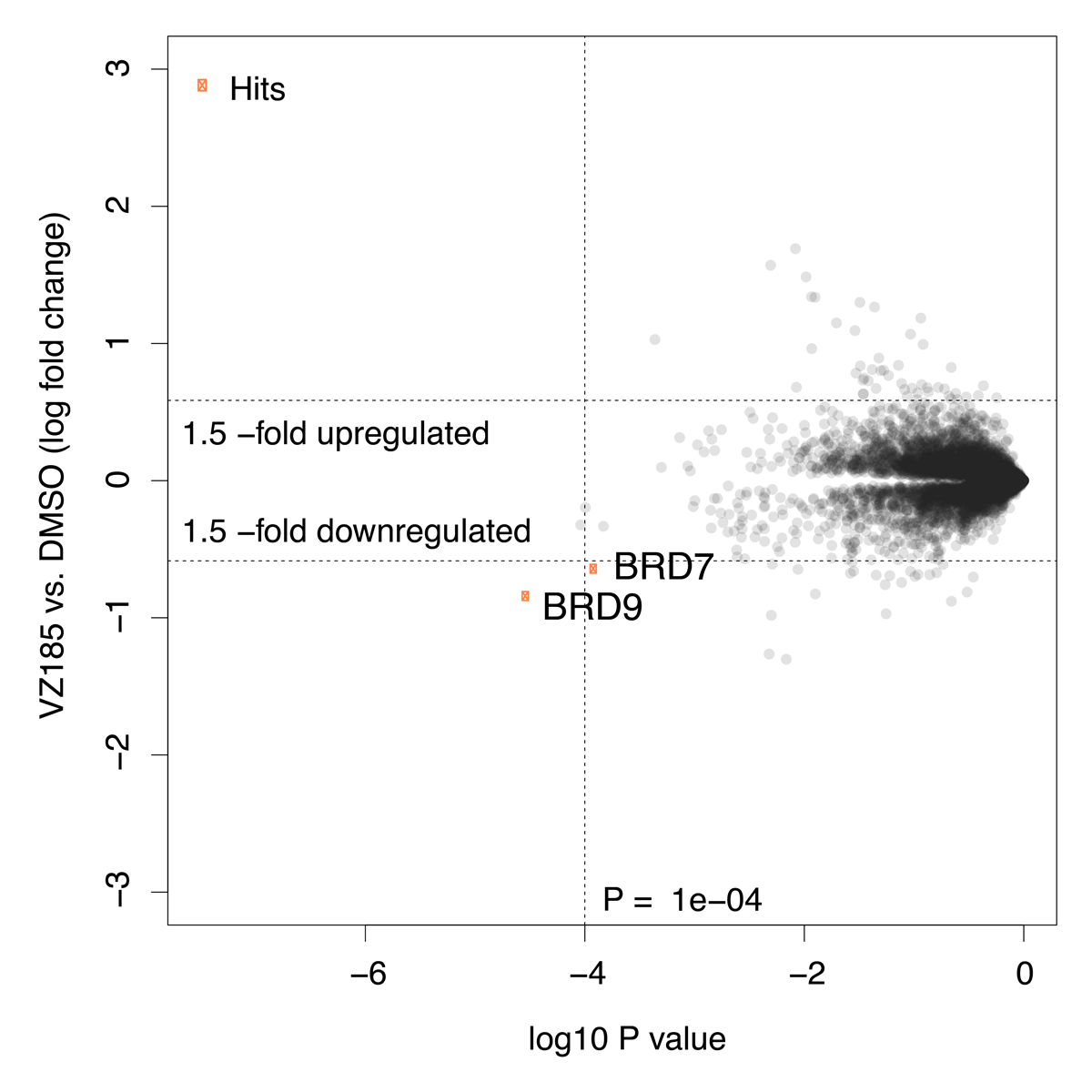
Global protein quantification was used to explore the unbiased proteome-wide selectivity of VZ185 induced degradation. Whole cell protein quantification was performed using label free quantification with the Fischer lab’s diaPASEF workflow.
As expected, BRD7/9 proteins were not depleted by treatment with negative control cis VZ185. Protein levels of other bromodomain-containing proteins or other BAF/PBAF subunits remained unaffected. To confirm selectivity over key potential off-target proteins within the bromodomain protein family, live cell kinetic analyses of endogenously tagged BRD2/3/4 and SMARCA4 proteins expressing LgBiT were performed, which showed no degradation of these proteins in the presence of VZ185. Together these results confirmed VZ185 as an effective and highly selective degrader of BRD7/9 proteins in cells1.
The PROTAC VZ185 was tested by the Eric Fisher Laboratory - Dana-Farber Cancer Institute as part of their Degradation Proteomics Initiative23,24. It induces selective degradation of BRD9 and BRD7 after 5 h of treatment at 0.1 µM in the neuroblastoma cell line Kelly Cells.
Of the 7,742 proteins quantified in this experiment, only BRD9 and BRD7 were found to be significantly downregulated in response to VZ185 treatment. Statistical analysis was performed using a moderated t-test in Bioconductor’s limma package to generate hit lists containing log2 Fold Change and P-values for each protein. The data are also displayed in the scatterplot above.
Download selectivity data:
VZ185_selectivityData.xlsx
cis_VZ185_selectivityData.xlsx
Degradation_Proteomics_Initiative_data_VZ185-BRD7-BRD9-PROTAC_1.xlsx
VZ185 is a VHL-based, potent and selective PROTAC (proteolysis-targeting chimera) degrader of the BAF/PBAF complexes subunits BRD7 & BRD9
{kind=link}
{kind=link}
Iterative design and optimization of initially inactive Proteolysis Targeting Chimeras (PROTACs) identify VZ185 as a potent, fast and selective von Hippel-Lindau (VHL)-based dual degrader probe of BRD9 and BRD7.
Discovery of I-BRD9, a Selective Cell Active Chemical Probe for Bromodomain Containing Protein 9 Inhibition
Theodoulou N. H., Bamborough P., Bannister A. J., Becher I., Bit R. A., Che K. H., Chung C. W., Dittmann A., Drewes G., Drewry D. H., Gordon L., Grandi P., Leveridge M., Lindon M., Michon A.-M., Molnar J., Robson S. C., Tomkinson N. C. O., Kouzarides T., Prinjha R. K., Humphreys P. G.
J. Med. Chem. 2016, 59, 1425–1439.
Integrative Genomics Analysis of Chromosome 5p Gain in Cervical Cancer Reveals Target Over-Expressed Genes, Including Drosha.
BRD7 Inhibits the Warburg Effect and Tumor Progression Through Inactivation of HIF1α/LDHA Axis in Breast Cancer.
A Major Chromatin Regulator Determines Resistance of Tumor Cells to T Cell–Mediated Killing.
LP99: Discovery and Synthesis of the First Selective BRD7/9 Bromodomain Inhibitor
Clark P. G. K., Vieira L. C. C., Tallant C., Fedorov O., Singleton D. C., Rogers C. M., Monteiro O. P., Bennett J. M., Baronio R., Müller S., Daniels D. L., Méndez J., Prof. Knapp S., Brennan P. E., Dixon D. J.
Angew. Chem. Int. Ed. Engl. 2015, 54, 6217-21.
Design and synthesis of potent and selective inhibitors of BRD7 and BRD9 bromodomains
Hay D. A., Rogers C. M., Fedorov O., Tallant C., Martin S., Monteiro O. P., Müller S., Knapp S., Schofielda C. J., Brennan P. E.
Med. Chem. Commun. 2016, 6, 1381-1386.
Structure-based design of an in vivo active selective BRD9 inhibitor
Martin L. J., Koegl M., Bader G., Cockcroft X.-L., Fedorov O., Fiegen D., Gerstberger T., Hofmann M. H., Hohmann A. F., Kessler D., Knapp S., Knesl P., Kornigg S., Müller S., Nar H., Rogers C., Rumpel K., Schaaf O., Steurer S., Tallant C., Vakoc C. R., Zeeb M., Zoephel A,, Pearson M., Boehmelt G., McConnell D.
J. Med. Chem. 2016, 59, 4462−4475.
Inhibition of Bromodomain-Containing Protein 9 for the Prevention of Epigenetically-Defined Drug Resistance.
Sensitivity and Engineered Resistance of Myeloid Leukemia Cells to BRD9 Inhibition.
The data can be accessed via the following link:
Mapping the Degradable Kinome Provides a Resource for Expedited Degrader Development
Donovan K. A., Ferguson F. M., Bushman J. W., Eleuteri N. A., Bhunia D., Ryu S., Tan L., Shi K., Yue H., Liu X., Dobrovolsky D., Jiang B., Wang J., Hao M., You I., Teng M., Liang Y., Hatcher J., Li Z., Manz T. D., Groendyke B., Hu W., Nam Y., Sengupta S., Cho H., Shin I., Agius M. P., Ghobrial I. M., Ma M. W., Che J., Buhrlage S. J., Sim T., Gray N. S., Fischer E. S.
Cell. 2020, 183(6):1714-1731.
When you plan a publication, please use the following acknowledgement:
VZ185 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.