Cereblon (CRBN) binder
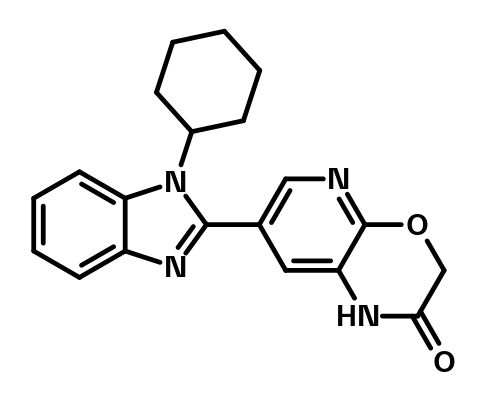
BI-3757
Cereblon (CRBN) binders applied for targeted protein degradation stabilize the closed conformation of CRBN. This conformation is presumed to be required for ubiquitinylation and subsequent degradation of the target protein. BI-3757 is a CRBN binder - jointly designed by the University of Dundee and Boehringer Ingelheim - which stabilizes the open-conformation. It thus serves as a useful tool to investigate the biological functions of CRBN conformation and its structural biology.
 binder | BI-3757")
 binder | BI-3757")
 binder | BI-3757")
More information
Cereblon (CRBN) is a crucial component of the CRL4 E3 ubiquitin ligase complex, playing a significant role in protein degradation. CRBN's function is highly influenced by its ability to switch between open and closed conformations. In its open conformation, CRBN is less effective at binding neosubstrates, which can be proteins targeted for degradation. However, when CRBN adopts its closed conformation, it can effectively bind and facilitate the ubiquitination of these neosubstrates, leading to their subsequent degradation by the proteasome1. This conformational change is often induced by CRBN binders, which enhance CRBN's ability to recruit and degrade specific proteins1,2. This mechanism has drawn a lot of attention, especially after it could be shown that the therapeutic mode of action of lenalidomide is based on its ability to harness the E3 ubiquitin ligase cereblon to selectively degrade its targets, the lymphoid transcription factors IKZF1 and IKZF3, and thus can be described as a CRBN binder3,4,5.
BI-3757 is a novel CRBN binder, which in contrast to the “classical thalidomide - lenalidomide-based ligands”, stabilizes the open-conformation. BI-3757 binds to the orthosteric tri-tryptophan pocket in the thalidomide-binding domain but is unable to close CRBN. It could therefore be utilized as chemical tool to explore biological and structural aspects of open vs. closed CRBN, and to probe the functional consequences of inhibiting CRBN without any confounding effects from CRBN-mediated degradation of neo-substrates1.
The molecule is the result of a collaboration between the University of Dundee, School of Life Sciences, and Boehringer Ingelheim.
To further stimulate research in this emerging area, we make BI-3757 and its negative control available for independent research.
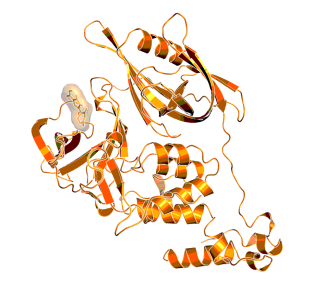
X-ray structure of the complex of CRBN with lenalidomide (PDB code: 5FQD), indicating the binding location of a small molecule “degrader”.
BI‑3757 shows biophysical binding to CRBN as measured by surface plasmon resonance (SPR) and cellular binding as assessed by NanoBRET.
| Probe name / Negative control | BI-3757 | BI-4506 |
| MW [Da]a | 348.4 | 362.4 |
| (KD) [nM]b | 217 | >10,000 |
| (IC50) [nM]c | 1,752 | >10,000 |
a The molecule is supplied in salt form; for the molecular weight of the salt, please refer to the vial label.
b Surface plasmon resonance (SPR): The CRBN target protein was biotinylated via its Avi‑tag and immobilized on a streptavidin sensor chip. Binding experiments were performed at 25 °C using a multi‑cycle approach with increasing molecule concentrations. Samples were injected over the immobilized protein, with dissociation times adjusted according to the individual off‑rates. Sensorgrams were reference‑ and blank‑corrected, and binding affinities (KD values) were determined using a steady‑state model, as all molecules showed fast binding kinetics.
c NanoBRET™ Target Engagement assay: This assay is used to assess CRBN binding and cellular permeability of CRBN‑based PROTACs in living cells. Experiments are performed in intact HEK293 cells expressing a NanoLuc‑CRBN fusion protein, with a parallel format in permeabilized cells to disentangle target binding from cell permeability effects. PROTAC binding to CRBN competes with a fluorescent CRBN tracer, resulting in tracer displacement and a corresponding decrease in the NanoBRET signal. Cells are incubated with test molecules over a concentration range, followed by addition of NanoLuc substrate and extracellular inhibitor, and BRET signals are measured at two wavelengths. Data are normalized to controls and fitted to dose–response curves to derive potency parameters, providing a quantitative measure of intracellular CRBN engagement and permeability.
BI‑3757 is a tool suitable for studying the effects of open and closed CRBN conformations in vitro. Due to low hepatic stability in rodents, in vivo use is not recommended.
| Probe name / Negative control | BI-3757 | BI-4506 |
| clogD @ pH 7.4 | 4.3 | 4.1 |
| Solubility @ pH 7 [µg/mL] | <1 | 47 |
| Caco-2 permeability AB @ pH 7.4 [*10-6 cm/s] | 69 | 76 |
| Caco-2 efflux ratio | 0.9 | 0.9 |
| MDCK permeability PappAB @ 1µM [10-6 cm/s] | 9.3 | 19.1 |
| MDCK efflux ratio | 0.9 | 0.7 |
| Microsomal stability (human/mouse/rat) [% QH] | 26 / 46 / 48 | <23 / 83 / 57 |
| Hepatocyte stability (human/mouse/rat) [% QH] | 21 / 85 / 81 | 14 / 96 / 86 |
| Plasma Protein Binding (human/mouse/rat) [%] | 94.6 / 97.7 / 92.3 | 95.1 / 96.8 / 92.0 |
| CYP 3A4 (IC50) [µM] | >50 | >50 |
| CYP 2C8 (IC50) [µM] | >50 | >50 |
| CYP 2C9 (IC50) [µM] | 47.7 | 19.1 |
| CYP 2C19 (IC50) [µM] | >50 | >50 |
| CYP 2D6 (IC50) [µM] | >50 | >50 |
| CYP 1A2 (IC50) [µM] | >50 | >50 |
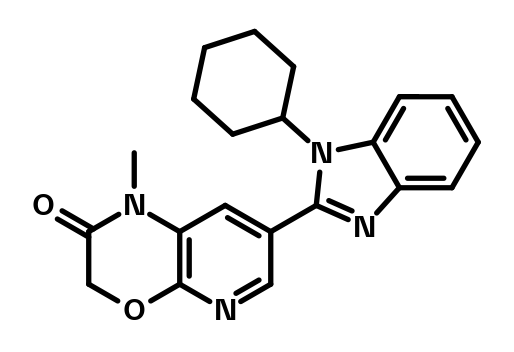
BI‑4506 is a structurally close, N‑methylated analog that does not bind CRBN and can be used as a negative control.
BI-4506 which serves as a negative control.
BI‑3757 inhibited 4 out of 44 targets tested by ≥ 50% at 10 µM (BZD/CENTR/R, 5‑HT2B/H AG, M2/H, and KAPPA [KOP]).
BI‑4506 inhibited 2 out of 44 targets tested by ≥ 50% at 10 µM (KAPPA [KOP] and M2/H).
| SELECTIVITY DATA AVAILABLE | BI-3757 | BI-4506 |
| SafetyScreen44™ with kind support of | Yes | Yes |
Download selectivity data:
BI-3757_selectivitydata.xlsx
BI-4506_selectivitydata.xlsx
The commercially available tool molecules lenalidomide, pomalidomide, and avadomide can serve as reference molecules amongst others.
{kind=link}
{kind=link}
Tuning the open-close equilibrium of Cereblon with small molecules influences protein degradation
O'Connor S., Rutter Z. J., Cowan A. D., Zeeb M., Binder F., Cao Y., Chakraborti S., Djukic S., Geist L., Hogg E., Kidd G., Krumb M., Langer S., Schmidt D., Martin L., McCrory E. H., Padroni G., Puoti I., Simpson L., Sturbaut M., Crozier L., Vetma V., Zhang Q., McAulay K., Theis T., Ciulli A.
BioRxiv 2025
Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells
4. Krönke J., Udeshi N. D., Narla A., Grauman P., Hurst S. N., McConkey M., Svinkina T., Heckl D., Comer E., Li X., Ciarlo C., Hartman E., Munshi N., Schenone M., Schreiber S. L., Carr S. A., Ebert B. L.
Science 2014, 343(6168), 301–305
When you plan a publication, please use the following acknowledgement:
BI-3757 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.