HSD17B13 inhibitor
BI-3231
BI-3231 is a highly potent inhibitor of human and mouse HSD17B13 with good selectivity against other HSD17B family members, such as HSD17B11. This compound has been extensively characterized both in vitro and in vivo and has shown good water solubility and permeability, as well as medium metabolic stability in human and mouse hepatocytes. Thus, it may be an excellent tool to explore the unknown biological function of HSD17B13.




More information
Hydroxysteroid 17-beta dehydrogenase 13 (HSD17B13) is a lipid droplet-associated member of the 17-beta hydroxysteroid dehydrogenases (HSD17B) family, primarily expressed in hepatocytes2.
Genome wide association studies (GWAS) revealed a strong genetic link between HSD17B13 and liver diseases, such as non-alcoholic fatty liver disease (NAFLD),3,4 non-alcoholic steatohepatitis (NASH) and cirrhosis5-8.
Although its biological function and endogenous substrates are unknown, HSD17B13 acts on a broad range of lipid substrates,9 including estradiol5. NAD+ (nicotinamide adenine dinucleotide, oxidized form) plays an important role as co-factor1,5.
Sequence similarities between HSD17B13 and other HSD17B isoforms indicate HSD17B11 as the closest homolog (85% sequence similarity based on MOE10 alignment), and thus was used to assess the selectivity of BI-3231. The overall sequence identity of murine and human HSD17B13 is 75% (source www.uniprot.org), displaying a higher conservation of 92% in the putative binding sites of NAD+ and BI-3231 based on computational modelling approaches1.
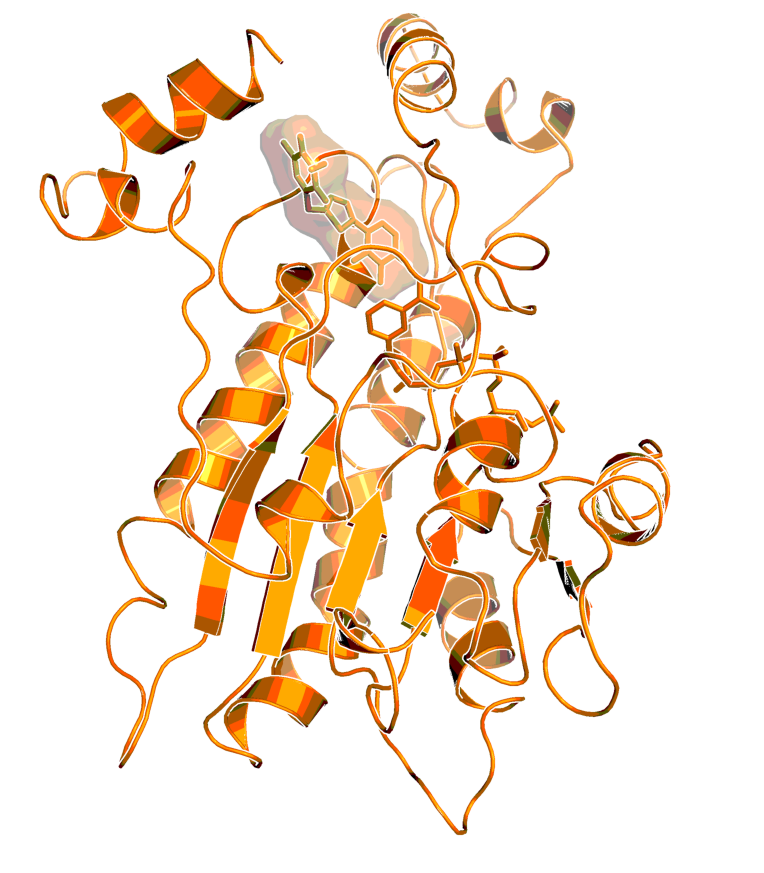
Binding model of BI-3231 in HSD17B131. BI-3231 is shown with molecular surface, the co-factor is shown without surface in stick representation.
BI-3231 shows low nanomolar potency in human and mouse HSD17B13 in vitro assays and good selectivity against HSD17B11. On-target binding was confirmed by a strong temperature shift in a human HSD17B13 thermal shift assay11. Interestingly, binding of BI-3231 to HSD17B13 only takes place in the presence of NAD+ (see Figure 4). These results indicate that binding of NAD+ is a prerequisite for the binding of BI-3231. Furthermore, the mode of inhibition was investigated via cross-titrations of BI-3231 and NAD+ revealing an uncompetitive mode of inhibition of BI-3231 against NAD+ (see graph below)1,12,13,14.
| Probe name / negative control | BI-3231 | BI-0955 |
| MW [Da, free base]a | 380.4 | 394.4 |
| Enzymatic hHSD17B13 assay (IC50 / Ki) [nM]b | (1)* / 0.7 | >10,000 |
| Cellular hHSD17B13 assay (IC50) [nM]b | 11 | >10,000 |
| Enzymatic hHSD17B11 assay (IC50) [nM]b | >10,000 | >10,000 |
| Enzymatic mHSD17B13 assay (IC50 / Ki) [nM]b | (14)* / 0.5 | >10,000 |
| DSF hHSD17B13 (temperature shift) [K]b | 16.7 | n. d. |
a For the salt form you will get, please refer to the label on the vial and for the molecular weight of the salt, please refer to the FAQs
b IC50 and Ki values are geometric means of multiple independent measurements (nd = not determined). *Real IC50 value unclear due to limits of the assay wall; Ki values (NAD+) should be used for comparison. For more detailed assay conditions please see literature.1
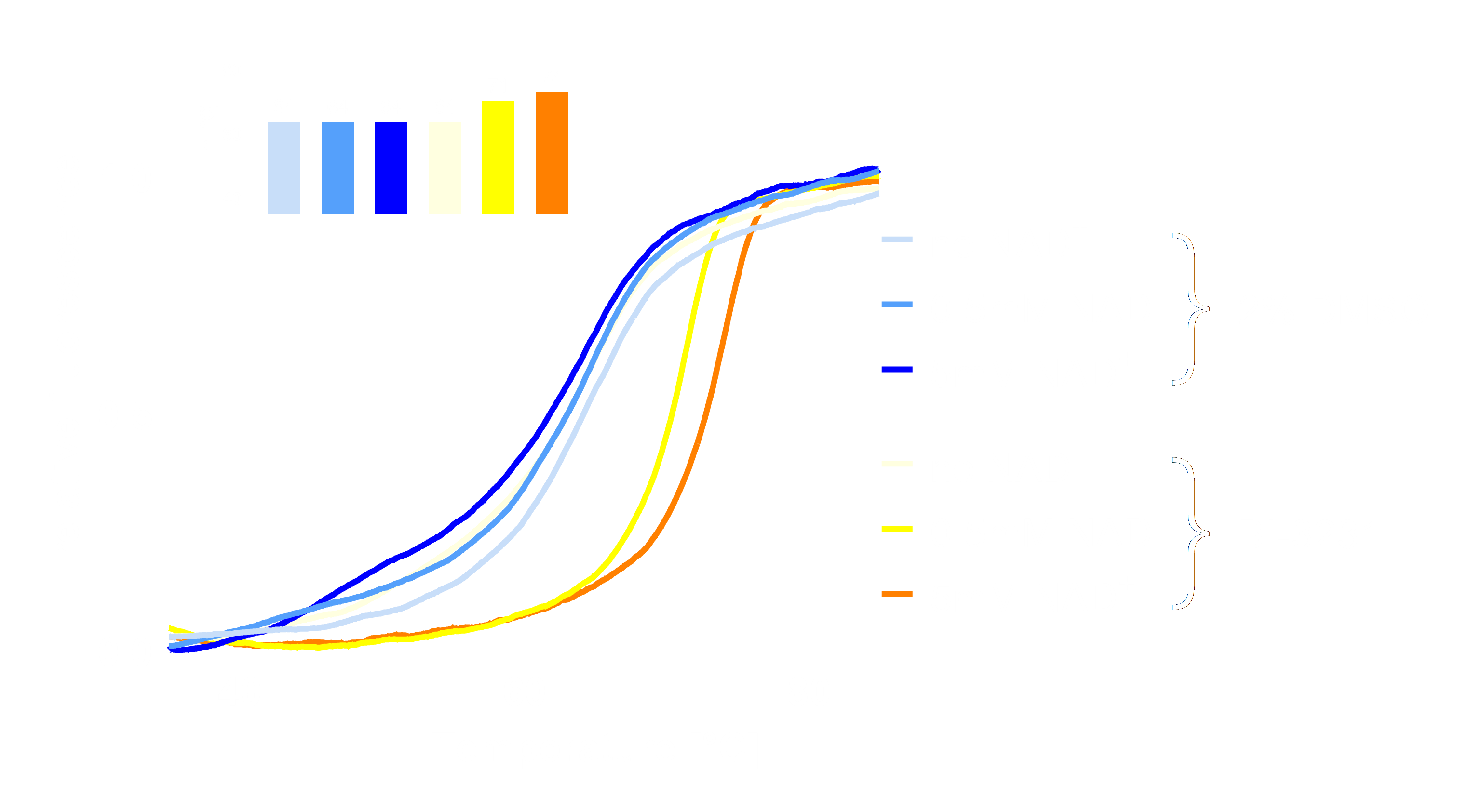
NAD+ dependency of BI-3231 binding: hHSD17B13 melting curves from Thermal Shift Assay experiment (nanoDSF) in the presence of 2% DMSO or BI-3231 at increasing NAD+ concentrations (0 mM, 0.5 mM and 5 mM) showed thermal stabilization by BI-3231 only in the presence of NAD+. Inset: corresponding melting temperatures.
Cross-titrations of BI-3231 at various NAD+ concentrations indicate an uncompetitive mode of inhibition of BI-3231 against NAD+.
BI-3231 shows good water solubility and permeability as well as medium metabolic stability in human and mouse hepatocytes1.
| Probe name / negative control | BI-3231 | BI-0955 |
| logD @ pH 2 / 11 | -1.2 | 3.0 |
| Solubility @ pH 6.8 [µg/mL] | >81 | <1 |
| Caco-2 permeability AB @ pH 7.4 [*10-6 cm/s] | 18 | 83 |
| Caco-2 efflux ratio | 1.2 | 1 |
| Microsomal stability (human/mouse) [% QH] | <23 / 25 | 59 / >88 |
| Hepatocyte stability (human/mouse) [% QH] | 58 / 57 | 68 / 97 |
| Plasma Protein Binding (human/mouse) [%] | 90.9 / 87.1 | 96.8 / 91.9 |
| hERG [inh. % @ 10 µM] | >10 | n. d. |
| CYP 3A4 (IC50) [µM] | >50 | >50 |
| CYP 2C8 (IC50) [µM] | >50 | >50 |
| CYP 2C9 (IC50) [µM] | >50 | >50 |
| CYP 2C19 (IC50) [µM] | >50 | 45.8 |
| CYP 2D6 (IC50) [µM] | >50 | >50 |
BI-3231 was thoroughly profiled in vivo (rodent PK, tissue distribution & bile excretion)1. Given the high clearance and short half-life of BI-3231, a tailored approach such as multiple daily administrations or the development of an extended-release formulation might be needed to maintain relevant target exposure in subchronic animal models.
| Probe name | BI-3231 |
| Clearance a [% QH] | 126.7 |
| Vss a [L/kg] | 1.4 |
| Mean residence time after i.v. dose [h]a | 0.2 |
| tmax [h]b | 0.25 |
| F [%] | 10 |
a i.v. dose: 1.9 mg/kg
b p.o. dose: 19 mg/kg
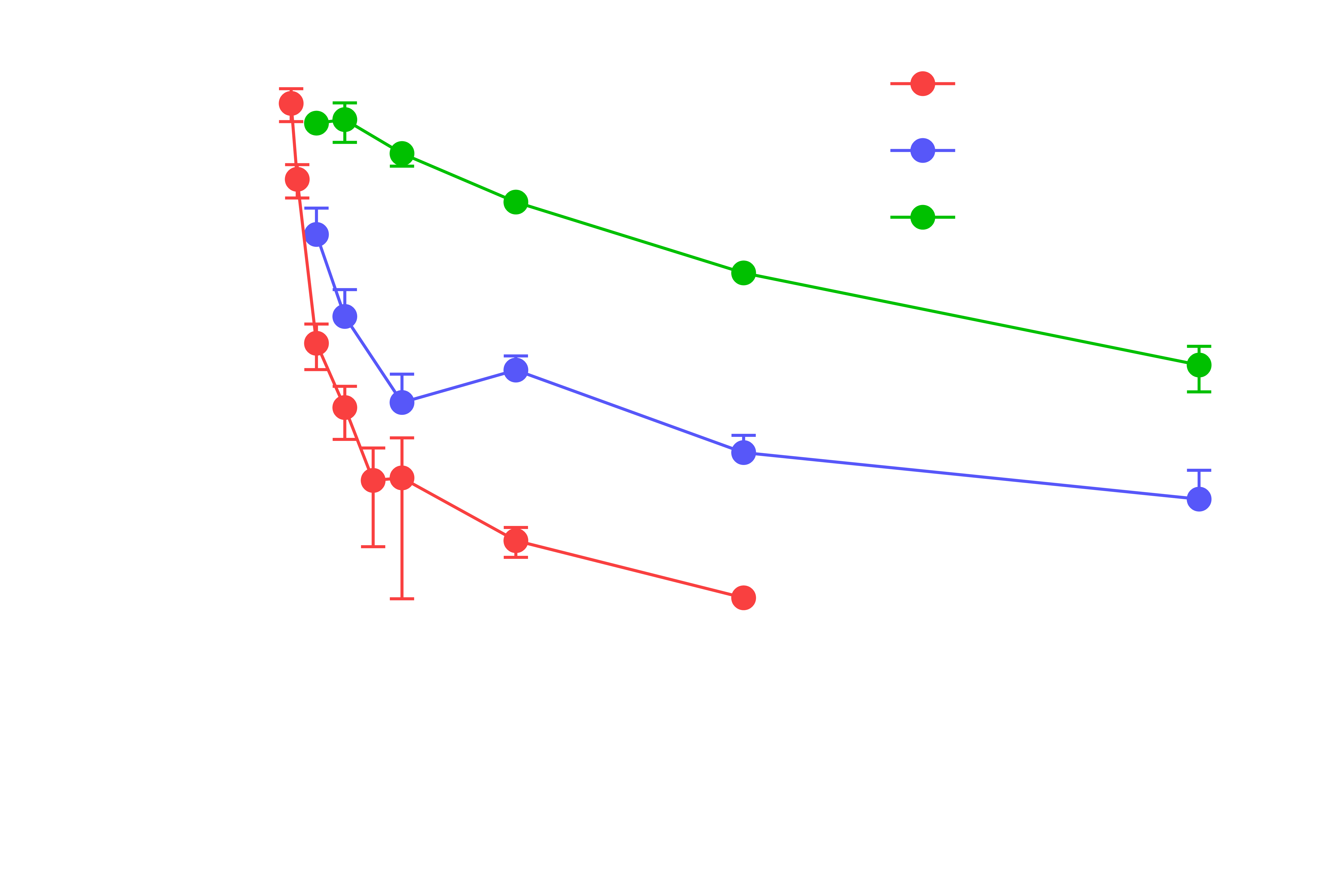
In vivo pharmacokinetics and tissue distribution of BI-3231 in mice (n = 3, SD indicated by error bars). The plasma pharmacokinetics after intravenous and oral administration in mice was characterized by a bi-phasic and rapid plasma clearance that exceeded the hepatic blood flow and a low oral bioavailability of 10%. Bioavailability was significantly increased by subcutaneous dosing. Relevant systemic exposure corresponding to >10-fold in vitro mouse Ki in unbound plasma concentration could be maintained over 8 hours in mice.
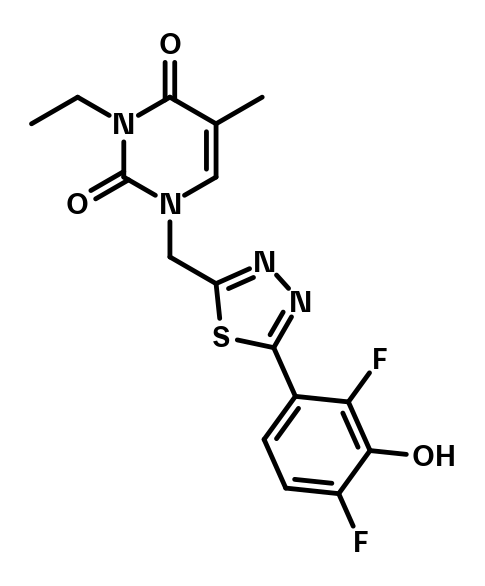
BI-0955 is the methylated analog of the active probe BI-3231. It does not have any detectable activity in the HSD17B13 in vitro assays.
BI-0955 serves as a negative control.
BI-3231 was tested against a panel of 44 receptors and showed an inhibition > 50% @ 10µM on 1 target (COX-2@CE). The negative control BI-0955 hit 2 targets (5HT2B/H and COX-2@CE).
| SELECTIVITY DATA AVILABLE | BI-3231 | BI-0955 |
| SafetyScreen44™ with kind support of | Yes | Yes |
| Invitrogen® | No | No |
| DiscoverX® | No | No |
| Dundee | No | No |
Download selectivity data:
BI-3231_selectivityData.xlsx
BI-0955_selectivityData.xlsx
BI-3231 is a highly potent and selective inhibitor of human and mouse of HSD17B13. BI-0955 is available as negative control.
{kind=link}
{kind=link}
Discovery of a novel potent and selective HSD17B13 Inhibitor BI-3231 a well-characterized Chemical Probe available for Open Science
Thamm S., Willwacher M. K., Aspnes G. E., Bretschneider T., Brown N. F., Buschbom-Helmke S., Fox T., Gargano E. M., Grabowski D., Hoenke C., Matera D., Mueck K., Peters S., Reindl S., Riether D., Schmid M., Tautermann C. S., Teitelbaum A. M., Trünkle C., Veser T., Winter M., Wortmann L.
J. Med. Chem. 2023 66(4):2832–2850.
Comparative Proteomic Study Reveals 17beta-HSD13 as a Pathogenic Protein in Nonalcoholic Fatty Liver Disease
Su W., Wang Y., Jia X., Wu W., Li L., Tian X., Li S., Wang C., Xu H., Cao J., Han Q., Xu S., Chen Y., Zhong Y., Zhang X., Liu P., Gustafsson J. A., Guan Y.
Proc National Acad Sci 2014, 111(31):11437-42.
A Protein-Truncating HSD17B13 Variant and Protection from Chronic Liver Disease
Abul-Husn N. S., Cheng X., Li A. H., Xin Y., Schurmann C., Stevis P., Liu Y., Kozlitina J., Stender S., Wood G. C., Stepanchick A. N., Still M. D., McCarthy S., O’Dushlaine C., Packer J. S., Balasubramanian S., Gosalia N., Esopi D., Kim S. Y., Mukherjee S., Lopez A. E., Fuller E. D., Penn J., Chu X., Luo J. Z., Mirshahi U. L., Carey D. J., Still C. D., Feldman M. D., Small A., Damrauer S. M., Rader D. J., Zambrowicz B., Olson W., Murphy A. J., Borecki I. B., Shuldiner A. R., Reid J. G., Overton J. D., Yancopoulos G. D., Hobbs H. H., Cohen J. C., Gottesman O., Teslovich T. M., Baras A., Mirshahi T., Gromada J., Dewey F. E.
N Engl J Med 2018, 378 (12), 1096–1106.
Hydroxysteroid 17- Beta Dehydrogenase 13 Variant Increases Phospholipids and Protects against Fibrosis in Nonalcoholic Fatty Liver Disease
Luukkonen P. K., Tukiainen T., Juuti A., Sammalkorpi H., Haridas P. A. N., Niemela O., Arola J., Orho-Melander M., Hakkarainen A., Kovanen P. T., Dwivedi O., Groop L., Hodson L., Gastaldelli A., Hyotylainen T., Oresic M., Yki-Jarvinen H.
Jci Insight 2020, 5 (5), e132158.
HSD17B13 Is a Hepatic Retinol Dehydrogenase Associated with Histological Features of Non-Alcoholic Fatty Liver
Ma Y., Belyaeva O. V., Brown P. M., Fujita K., Valles K., Karki S., Boer Y. S. de, Koh C., Chen Y., Du X., Handelman S. K., Chen V., Speliotes E. K., Nestlerode C., Thomas E., Kleiner D. E., Zmuda J. M., Sanyal A. J., Nash C. R. N., Kedishvili N. Y., Liang T. J., Rotman Y.
Hepatology 2019, 69 (4), 1504–1519.
Genetic Variation in HSD17B13 Reduces the Risk of Developing Cirrhosis and Hepatocellular Carcinoma in Alcohol Misusers
Stickel F., Lutz P., Buch S., Nischalke H. D., Silva I., Rausch V., Fischer J., Weiss K. H., Gotthardt D., Rosendahl J., Marot A., Elamly M., Krawczyk M., Casper M., Lammert F., Buckley T. W., McQuillin A., Spengler U., Eyer F., Vogel A., Marhenke S., Felden J. von, Wege H., Sharma R., Atkinson S., Franke A., Nehring S., Moser V., Schafmayer C., Spahr L., Lackner C., Stauber R. E., Canbay A., Link A., Valenti L., Grove J. I., Aithal G. P., Marquardt J. U., Fateen W., Zopf S., Dufour J. F., Trebicka J., Datz C., Deltenre P., Mueller S., Berg T., Hampe J., Morgan M. Y.
Hepatology 2020, 72 (1), 88–102.
17beta- Hydroxysteroid Dehydrogenases (17beta-HSDs) as Therapeutic Targets: Protein Structures, Functions, and Recent Progress in Inhibitor Development
Marchais-Oberwinkler S., Henn C., Moller G., Klein T., Negri M., Oster A., Spadaro A., Werth R., Wetzel M., Xu K., Frotscher M., Hartmann R. W., Adamski J.
J Steroid Biochem Mol Biol 2011, 125 (1–2), 66–82.
Molecular Operating Environment (MOE)
2022.02 Chemical Computing Group ULC, 1010 Sherbooke St. West, Suite #910, Montreal, QC, Canada, H3A 2R7, 2022.
Chemical Computing Group (CCG) | Computer-Aided Molecular Design (chemcomp.com)
When you plan a publication, please use the following acknowledgement:
BI-3231 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.