KHK inhibitor
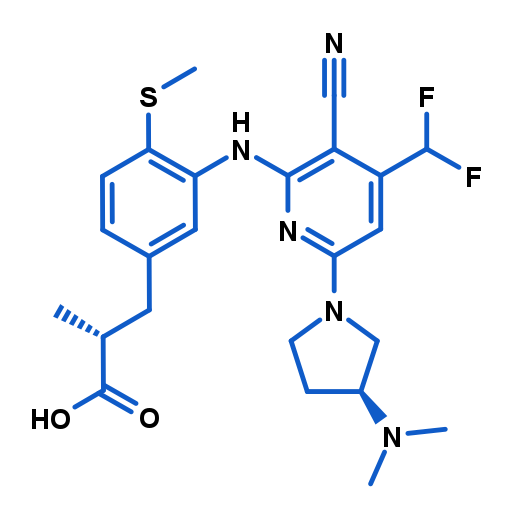
BI-9787
BI-9787 is a potent, structurally distinct, zwitterionic KHK inhibitor, suitable for use in vitro as well as in vivo. It combines high potency and high permeability with a favorable oral rat PK and high target selectivity. In addition, we also provide BI-2817, which can serve as a negative control for in vivo and in vitro studies that utilize BI-9787 as a chemical probe.



More information
Ketohexokinase (KHK) is a kinase that catalyzes the phosphorylation of fructose, forming fructose-1-phosphate. In humans, KHK exists as two isoforms, KHK-A and KHK-C, whereby the former has a lower affinity for fructose and is expressed in various tissues1,2. The higher fructose affinity KHK-C is expressed predominately in the liver, pancreas, kidneys, and small intestine1. Since fructose metabolism occurs mainly in the gut wall and, in instances of high fructose consumption, the liver, pancreas, and kidneys, KHK-C is thought to play a key role in metabolic disorders, implicating KHK-C as a driver of metabolic dysfunction2.
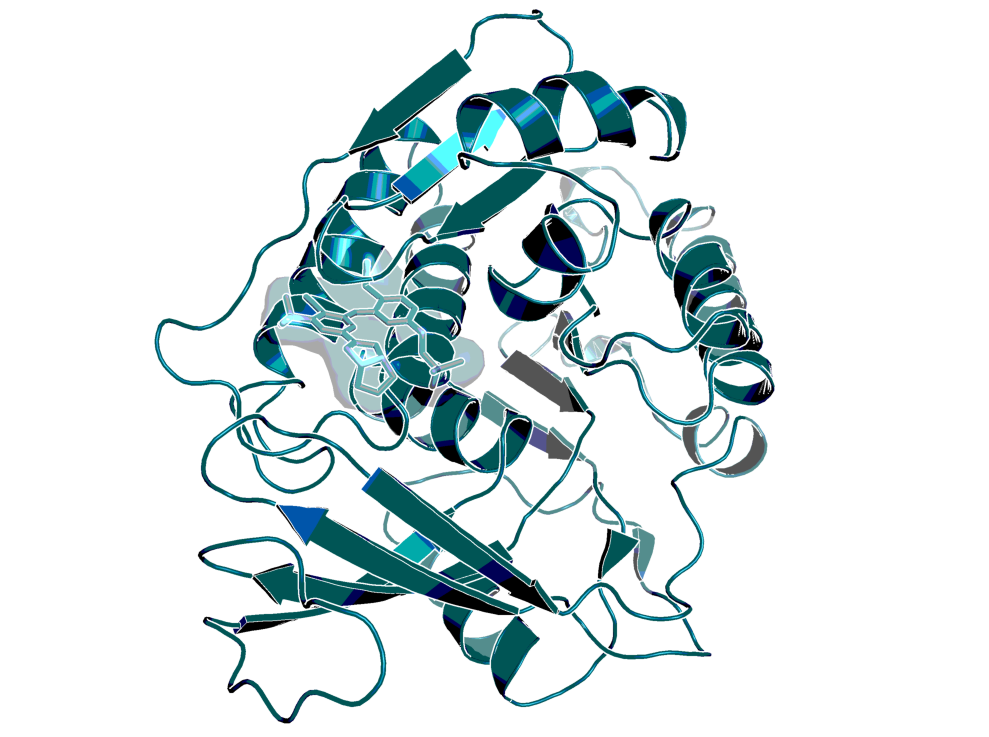
X-ray structure of a close BI-9787 analog in association with KHK-C (PDB code: 8OMJ).
The zwitterionic structure of BI-97873 displays a combination of high enzymatic and cellular potency, high microsomal stability and significantly improved selectivity.
| Probe name / Negative control | BI-9787 | BI-2817 |
| MW [Da, free base]a | 489.6 | 447.5 |
| hKHK-C (IC50) [nM]b | 12.8 | 5,029 |
| hKHK-A (IC50) [nM]b | 12 | n.a. |
| Mouse KHK-C (IC50) [nM]b | 20 | n.a. |
| Rat KHK-C (IC50) [nM]b | 3.0 | 8,870 |
| F1P HepG2 (IC50) [nM]c | 123 | 12,256 |
| F1P mouseHep (IC50) [nM]c | 59 | n.a. |
a For the salt form you will get, please refer to the label on the vial and for the molecular weight of the salt, please refer to the FAQs
bAssay conditions: Kinase activity of recombinant His-tagged KHK isoforms.
The enzymatic activity of recombinant human KHK-C, human KHK-A, rat and mouse KHK-C was determined using the ADP-GLOTM Kinase Assay kit from Promega as described in the instructions. In brief, 1.25 µg/ml His tagged human KHK-A was incubated for 60 min at room temperature with 15 mM D-fructose. His-tagged human KHK-C (1 µg/ml) and His-tagged mouse KHK-C (0.625 µg/ml) were incubated for 60 min at room temperature with 400 µM D-fructose and 200 µM ATP. His-tagged rat KHK-C (0.5 µg/1.5 ml) was incubated for 60 min at room temperature with 100 µM D-fructose and 200 µM ATP.
For all incubations the following assay buffer was used: 50mM HEPES pH 7.4, 4mM MgCl2, 20mM KCl, 0.01% Tween 20, 1mM DTT. The enzymatic reaction was stopped and developed using the ADP-GLOTM Kinase kit according to the manufacturers description and the results were analyzed using the luminescence signal determined by a multiplate reader from EnVision. Signals from samples with enzyme and substrates alone were reported as 100% and signals from samples with enzyme alone were reported as 0%.
cAssay conditions: Quantitative determination of fructose-1-phosphate in HepG2 cells and primary mouse hepatocytes.
HepG2 cells (B.B. Knowles, Wistar Institute) were incubated with test compound or solvent (DMSO) in medium (EMEM, 10 mM NEAA, 8 mM glutamine, 10% FCS) for 30 min at 37°C under 5 % CO2. D-fructose was added to a final concenItration of 15 mM and cells were incubated for a further 60 min under the same conditions. Cells were put on ice washed with phosphate buffered saline and lysed in 10 mM ammonium acetate. Cell protein was precipitated with acetonitrile and an aliquot of the supernatant was analyzed for fructose-1-phosphate using the RapidFire-MS/MS (RIAS) technology. Fructose-6-phosphate (0.1 µM) in the samples was used as internal standard for the quantification.
Primary mouse hepatocytes (Fisher Scientific) were incubated in Williams Medium E (0,1 mg/ml Gentamycin, 0.1 µM Dexamethasone, 2 mM L-glutamine, 17 nM insulin-transferrin-selenite supplement-G). All other reagents and procedures were identical with the protocol described for HepG2 cells.
BI-9787 and BI-2817 show good cell permeability, good hepatocyte stability across species and high plasma protein binding.
| Probe name / negative Control | BI-9787 | BI-2817 |
| logD @ pH 11 | 0.9 | 0.6 |
| High Throughput Solubility @ pH 6.8 [µg/mL] | 19 | 87 |
| Caco-2 permeability AB @ pH7.4 [*10-6 cm/s] | 22 | 19 |
| Caco-2 efflux ratio | 1.9 | 1.7 |
| Microsomal stability (human/mouse/rat) [% QH] | <23/<23/<22 | 25/<23/<22 |
| Hepatocyte stability (human/mouse/rat) [% QH] | 24 / 25 / 13 | 23 / 15 / 10 |
| Plasma Protein Binding (human/mouse/rat) [%] | >99.9 / 99.4 / 99.5 | 99.6 / 95.7 / 95.0 |
| hERG (IC50) [µM] | 1.6 | >10 |
| CYP 3A4 (IC50) [µM] | >50 | >50 |
| CYP 2C8 (IC50) [µM] | 26.8 | >50 |
| CYP 2C9 (IC50) [µM] | >50 | >50 |
| CYP 2C19 (IC50) [µM] | >50 | >50 |
| CYP 2D6 (IC50) [µM] | >50 | >50 |
BI-9787 exhibits good clearance and mean residence time in rat together with good bioavailability.
| BI-9787 | RAT |
| Clearence [% QH]a | 22 |
| Mean residence time after i.v. dose [h]a | 2 |
| tmax [h]b | 0.4 |
| Cmax [nM]b | 2,620 |
| F [%] | 68 |
| Vss [l/kg] | 1.8 |
a i.v. dose: 0.5 mg/kg
b p.o. dose: 5 mg/kg
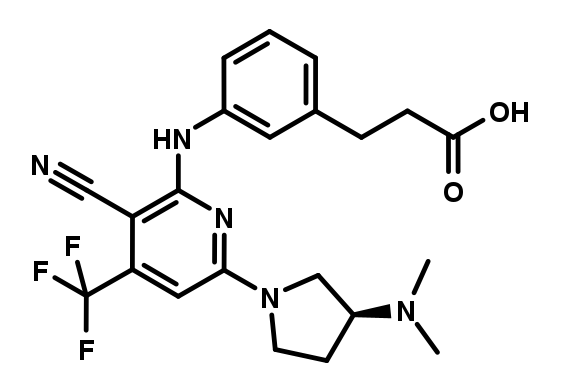
BI-2817 was designed as an inactive analogue of BI-9787, resulting in a tool compound that is structurally very close to BI-9787 and displays almost identical in vitro PK parameters.
BI-2817 which serves as a negative control.
BI-9787 was tested on 44 targets in a selectivity panel and showed ≥100 fold selectivity for 43 targets. Against PDE4D2, it still maintains an estimated selectivity factor of at least 30-fold (81% inhibition at 10 µM). The comparably selective negative control BI-2817 showed in only 3 out of 44 targets inhibition with more than 50% at 10 µM (N_Neuro_A, H1/Pyril/HS, BZD/Centr/R).
| Selectivity data available | BI-9787 | BI-2817 |
| SafetyScreen44™ with kind support of | Yes | Yes |
| Invitrogen® | No | No |
| DiscoverX® | No | No |
| Dundee | No | No |
Download selectivity data:
BI-9787_selectivityData.xlsx
BI-2817_selectivityData.xlsx
PF-068359194-7 can be used as a reference compound.
BI-9787 is a potent, highly permeable and selective KHK inhibitor, suitable for in vitro and in vivo use. The structurally similar molecule BI-2817 is provided as a negative control.
{kind=link}
{kind=link}
Opposing effects of fructokinase C and A isoforms on fructose-induced metabolic syndrome in mice
Ishimoto T., Lanaspa M. A., Le MyPhuong T., Garcia G. E., Diggle C. P., MacLean P. S., Jackman M. R., Asipu A., Roncal-Jimenez C. A., Kosugi T., Rivard C. J., Maruyama S., Rodriguez-Iturbe B., Sánchez-Lozada L. G., Bonthron D. T., Sautin Y. Y., Johnson R. J.
Proceedings of the National Academy of Sciences 2012, 109 (11), 4320-4325.
Pharmacologic inhibition of ketohexokinase prevents fructose-induced metabolic dysfunction
Gutierrez J. A., Liu W., Perez S., Xing G., Sonnenberg G., Kou K., Blatnik M., Allen R., Weng Y., Vera N. B., Chidsey K., Bergman A., Somayaji V., Crowley C., Clasquin M. F., Nigam A., Fulham M. A., Erion D. M., Ross T. T., Esler W. P., Magee T. V., Pfefferkorn J. A., Bence K. K., Birnbaum M. J., Tesz G. J.
Molecular Metabolism 2021, 48, 101196.
Discovery of PF-06835919: A Potent Inhibitor of Ketohexokinase (KHK) for the Treatment of Metabolic Disorders Driven by the Overconsumption of Fructose
Futatsugi K., Smith A. C., Tu M., Raymer B., Ahn K., Coffey S. B., Dowling M. S., Fernando D. P., Gutierrez J. A., Huard K., Jasti J., Kalgutkar A. S., Knafels J. D., Pandit J., Parris K. D., Perez S., Pfefferkorn J. A., Price D. A., Ryder T., Shavnya A., Stock I. A., Tsai A. S., Tesz G. J., Thuma B. A., Weng Y., Wisniewska H. M., Xing G., Zhou J., Magee T. V.
Journal of Medicinal Chemistry 2020, 63 (22), 13546-13560.
A phase 2a, randomized, double-blind, placebo-controlled, 3-arm, parallel-group study to assess the efficacy, safety, tolerability and pharmacodynamics of PF-06835919 in patients with nonalcoholic fatty liver disease and type 2 diabetes
Saxena A. R., Lyle S.-A., Khavandi K., Qiu R., Whitlock M., Esler W. P., Kim A. M.
Diabetes Obes Metab. 2023, 25(4):992-1001.
When you plan a publication, please use the following acknowledgement:
BI-9787 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.