Secretin Receptor Agonist
BI-3434
BI-3434 is a potent peptidic secretin receptor agonist that serves as a high-quality in vitro and in vivo tool compound1. The agonist shows high potency with good selectivity and a prolonged half-life in mice. Increased energy expenditure after daily subcutaneous treatment with BI-3434 suggests that secretin receptor may be involved in metabolic regulation and energy homeostasis.



More information
Secretin, a 27-amino acid peptide, is synthesized in the enteroendocrine S-cells located in the duodenum and proximal jejunum. It is produced in response to low pH (pH<4.5) and the uptake of fatty acids in the stomach2. Acting upon the secretin receptor (SctR), which belongs to the class B G-protein-coupled receptor (GPCR) family3, this peptide induces both peripheral and central effects4. First discovered in 1902, secretin is primarily recognized for its role in stimulating the secretion of pancreatic and biliary hydrogen carbonate and water in order to neutralize gastric acid within the duodenum. It also plays a crucial role in regulating pancreatic enzyme secretion5. Additionally, secretin has been found to inhibit gastric emptying6 and promote the secretion of pancreatic islet hormones7,8. Moreover, secretin exhibits potential for the treatment of obesity. Studies using SctR knock-out mice have revealed that these animals are resistant to obesity induced by a high-fat diet and demonstrate impaired lipid absorption9. Administration of secretin, either peripherally or centrally, in rodents has been found to reduce food intake10 and stimulate lipolysis11. Furthermore, continuous subcutaneous administration of secretin transiently elevates energy expenditure and reduces body weight12.
BI-3434 activated target tissues like pancreas, adipose tissue, and stomach in vivo. BI-3434 also increased energy expenditure after daily administration leading to a loss of fat mass, although this did not translate in a significant effect on body weight. However, treatment in combination with a GLP-1R agonist led to a synergistic effect on body weight loss. Details can be found in reference 1.
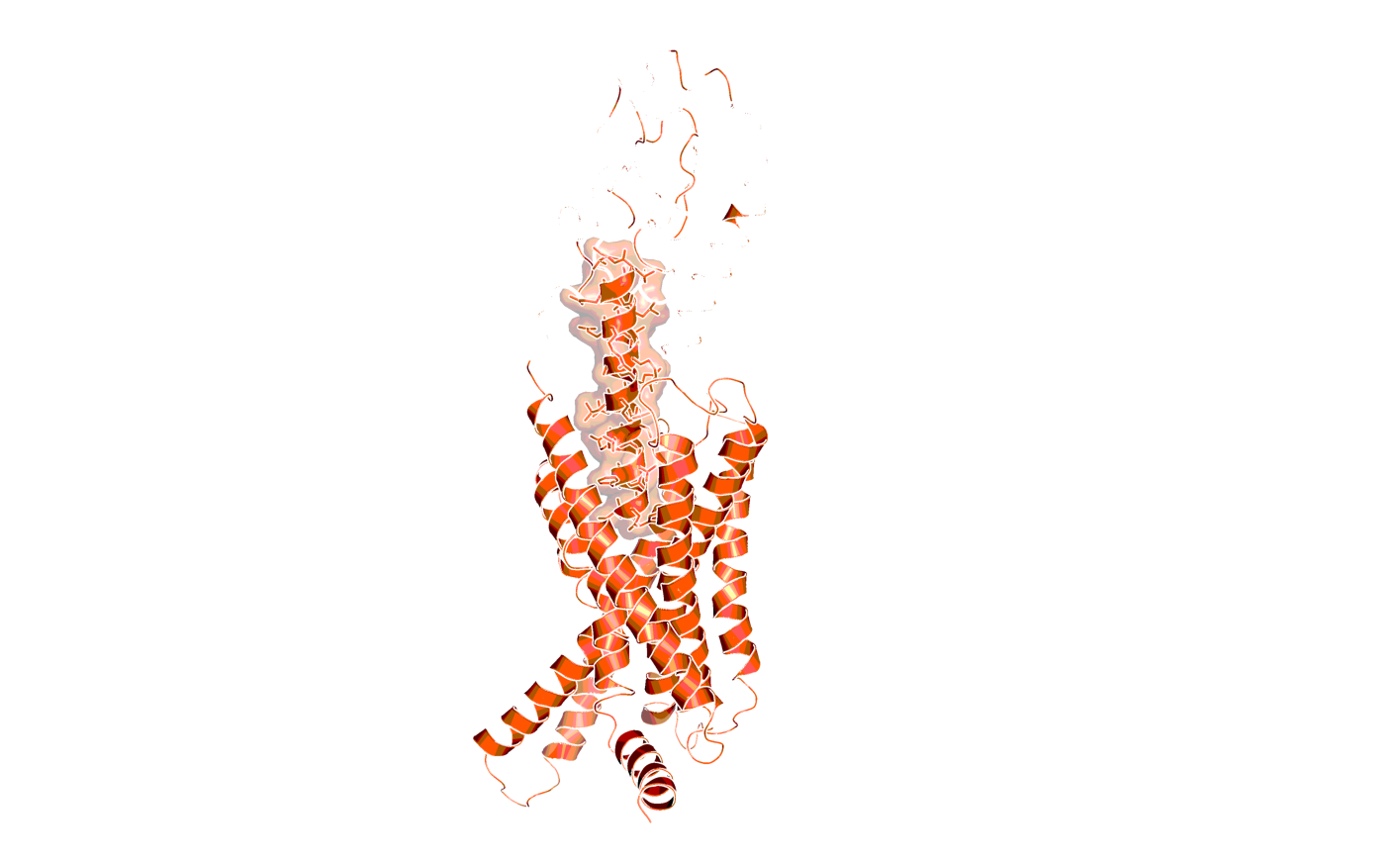
Structure of the Secretin receptor (magenta) in complex with its agonist secretin (helix depicted in white, atoms and surface shown; PDB code 6WZG)
| Probe name | BI-3434 |
| MW [Da, free base]a | 3938.6 |
| hSctR EC50 [pM]b | 15.5 |
| hGLP-1R in 0.5% hPlasma EC50 [nM]c | >100 |
| hGIPR in 0.5% hPlasma EC50 [nM]d | >100 |
| hGCGR in 0.5% hPlasma EC50 [nM]e | >990 |
a For the salt form you will get, please refer to the label on the vial and for the molecular weight of the salt, please refer to the FAQs
bAssay conditions: Functional cAMP assay with cAMP Hunter™ CHO-K1 SCTR Gs Cell Line
The in vitro functional cAMP assay was used to measure accumulation of cAMP after receptor activation in cells stably expressing human SCTR (cAMP Hunter™ CHO-K1 SCTR Gs Cell Line, Eurofins/DiscoverX). All cAMP assays were conducted at Eurofins/DiscoverX according to the following protocol. cAMP Hunter cell lines were expanded from freezer stocks according to standard procedures. Cells were seeded in a total volume of 20 μL into white walled, 384-well microplates and incubated at 37 °C for the appropriate time prior to testing. cAMP modulation was determined using the DiscoverX HitHunter cAMP XS+ assay. For agonist determination, cells were incubated with human secretin or with BI-3434 to induce response. Medium was aspirated from cells and replaced with 15 μL 2:1 HBSS/10mM Hepes: cAMP XS+ Ab reagent. Intermediate dilution of sample stocks was performed to generate 4x compound in assay buffer. 5 μL of 4x compound was added to cells and incubated at 37 °C or room temperature for 30 or 60 minutes. After appropriate compound incubation, assay signal was generated through incubation with 20 μL cAMP XS+ ED/CL lysis cocktail for 1h followed by incubation with 20 μL cAMP XS+ EA reagent for 3h at room temperature. Microplates were read following signal generation with a PerkinElmer EnvisionTM instrument for chemiluminescent signal detection. Compound activity was analyzed using CBIS data analysis suite (ChemInnovation, CA). For Gs agonist mode assays, percentage activity was calculated using the following formula: % Activity =100% x (mean RLU of test compound - mean RLU of vehicle control) / (mean RLU of MAX control - mean RLU of vehicle control).
bAssay conditions: Human GLP1R Cre-Luc Assay with 0.5% human plasma
Human HEK293 CRE-luc2P cells (Promega, Madison, WI, USA) expressing recombinant GLP-1R and GCGR were cultivated in Dulbecco’s Modified Eagle Medium (with high glucose/l-glutamine) supplemented with 10% fetal bovine serum, 50 μg/mL hygromycin, and 400 μg/mL Geneticin™. For the assays, cells were resuspended either in KRBH with 0.5% human or mouse plasma or in 100% human or mouse plasma. Cells were treated for 4 h at 37 °C with the different peptides (all n = 3; all tested peptides were produced in house, handled as 1 mM stock solution in dimethyl sulfoxide, and tested at a final concentration range between 0.2 pM and 1 μM). In vitro potency was assessed using the Bright-Glo™ Luciferase Assay System (Promega), measuring the production of cAMP through a CRE-controlled luciferase. The potency (EC50), plasma shift, and GLP-1R/GCGR ratio was calculated for each plasma condition.
cAssay conditions: Human GIPR Cre-Luc Assay with 0.5% human plasma
Stably hGIPR-transfected CHO cells (20,000 cells/well; 384-well format) are incubated with test peptides in the presence of 0.5% plasma for 4 hours at 37°C in a humidified incubator. Following incubation, assay plates are equilibrated to room temperature, followed by addition of an equal volume of Bright-Glo™ Luciferase reagent, incubation at room temperature for 10 minutes and analysis of luminescence (Envision Reader).
dAssay conditions: Human GCGR cAMP HTRF Assay with 0.5% human Plasma
The principle of the used cAMP HTRF assay kit is that a FRET signal is generated between the donor label of a tagged cAMP (cAMP-Eu) and the acceptor label of an anti-cAMP antibody (anti-cAMP-d2). Stimulation of hGCGR with an agonistic peptide increases intracellular cAMP levels, which in turn leads to a reduction of the FRET signal.
Stably hGCGR-transfected HEK293 cells (1,000 cells/well; 384-well format) are incubated with test peptides in the presence of 0.5% plasma for 40 minutes at 37°C in a humidified incubator. Following incubation, assay plates are equilibrated to room temperature, followed by addition of both FRET reagents, incubation at room temperature for 1 hour and measurement of the FRET signal (Envision Reader).
BI-3434 shows good solubility and physical stability at pH 4.
| Probe name | BI-3434 |
| logD @ pH 11 | 1.5 |
| Isoelectric point | 6.9 |
| Solubility @ pH 4.0/pH 7.0 [mg/mL] | 3000 / <1 |
| Physical stability in THT assay @ pH 4 | Stable |
| Caco-2 permeability AB @pH7.4 [*10-6 cm/s] | <0.1 |
BI-3434 shows prolonged mean residence time in mice.
| BI-3434 | Mouse |
| Clearance [% QH]a | 0.6 |
| Mean residence time after i.v. dose [h]a | 5.7 |
| tmax [h]b | 2 |
| Cmax [nM]b | 70.9 |
| F [%] | 62 |
| Vss [L/kg]a | 0.19 |
a i.v. dose: 0.11 mg/kg
b s.c. dose: 0.11 mg/kg
BI-3434 was tested on 44 targets in a selectivity panel and showed selectivity for 42 targets (≤ 50% inhibition @ 10 µM). In two assays (COX-2 and LCK_CE) the compound showed inhibition between 70-83% @ 10µM. Both targets are intracellular and not accessible by the lipidated peptide BI-3434.
| SELECTIVITY DATA AVILABLE | BI-3434 |
| SafetyScreen44™ with kind support of | Yes |
| Invitrogen® | No |
| DiscoverX® | No |
| Dundee | No |
Download selectivity data:
BI-3434_selectivityData.xlsx
BI-3434 is a potent, selective and long-acting peptidic secretin receptor agonist, suitable for in vitro and in vivo studies, with prolonged half-life.
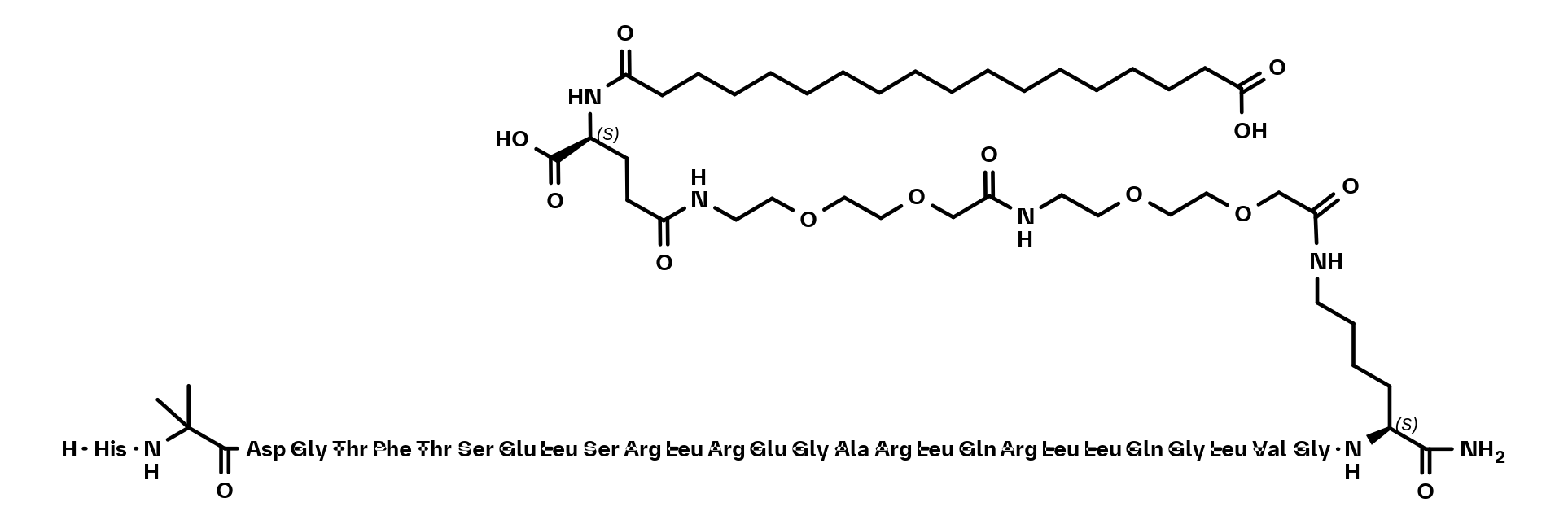
2D structure formats available
Secretin Receptor Agonist | BI-3434.png
{kind=link}
Receptors for secretin, calcitonin, parathyroid hormone (PTH)/PTH-related peptide, vasoactive intestinal peptide, glucagonlike peptide 1, growth hormone-releasing hormone, and glucagon belong to a newly discovered G-protein-linked receptor family
Segre G. V., Goldring S. R.
Trends Endocrinol Metabolism 1993, 4(10):309-14.
EFFECT OF SECRETIN ON INSULIN SECRETION
Unger R. H., Ketterer H., Eisentraut A., Dupré J.
Lancet 1966, 288, 24–26.
Secretin-Activated Brown Fat Mediates Prandial Thermogenesis to Induce Satiation
Li Y., Schnabl K., Gabler S.-M., Willershäuser M., Reber J., Karlas A., Laurila S., Lahesmaa M., Din M. U., Bast-Habersbrunner A., Virtanen K. A., Fromme T., Bolze F., O’Farrell L. S., Alsina-Fernandez J., Coskun T., Ntziachristos V., Nuutila P., Klingenspor M.
Cell 2018, 175(6):1561-1574.e12.
When you plan a publication, please use the following acknowledgement:
BI-3434 was kindly provided by Boehringer Ingelheim via its open innovation platform opnMe, available at https://www.opnme.com.